A1 Skorpione
Hinweis: In der Prüfung musst du von den Aufgabenblöcken A, B und C jeweils einen der beiden Vorschlägen 1 oder 2 bearbeiten.
Skorpione (Scorpiones) sind eine Ordnung der Spinnentiere (Arachnida), deren charakteristische Merkmale zum einen der Giftstachel am Ende des Hinterleibs und zum anderen die Fangarme mit Scheren sind. Von den ungefähr 2000 weltweit bekannten Arten gelten etwa 25 aufgrund ihres Giftes für den Menschen als potenziell tödlich.
Im Gift des Gelben Fünfstreifenskorpions (Leiurus quinquestriatus) kommen u. a. die in Tabelle 1 aufgeführten Nervengifte vor, mit deren Hilfe er z. B. Mäuse erbeutet.
Tab. 1: Auswahl von Nervengiften des Gelben Fünfstreifenskorpions1
| Nervengift | Wirkung und Wirkort |
|---|---|
| α-Toxin | Daueröffnung von postsynaptischen Natriumionen-Kanälen |
| Agitoxin 1 | Hemmung von Kaliumionen-Kanälen in der Axonmembran |
Fertige eine beschriftete Skizze einer chemischen Synapse im Nervensystem eines Beutetiers an.
Zeichne in Abbildung 1 den Verlauf eines postsynaptischen Potentials ein, das sich bei einer Gabe von α-Toxin zum Zeitpunkt 0 ms ergibt, und begründe den Unterschied in den Kurvenverläufen mit und ohne Giftgabe auf molekularer Ebene.
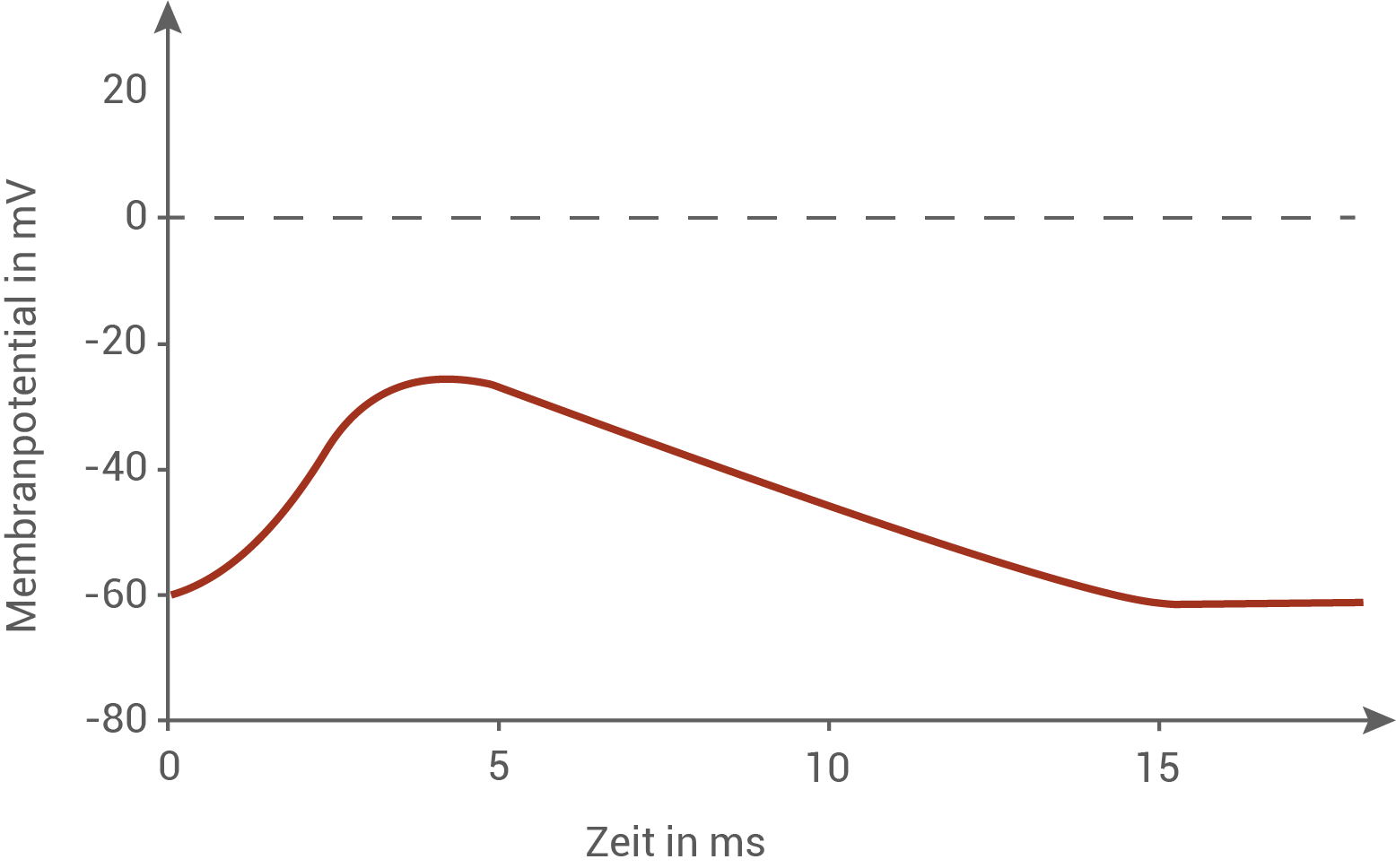
Erläutere die Wirkung von Agitoxin 1 auf die Erregung am Axon eines Beutetiers unter Berücksichtigung der Ionentheorie.
Um die Giftigkeit eines Skorpions einzuschätzen, gibt es eine weit verbreitete Faustregel: Je kleiner ein Skorpion ist, umso stärker ist sein Gift. Es wird zudem diskutiert, ob man statt der Körperlänge auch das Dickenverhältnis zwischen Scheren und Schwanz zur Einschätzung der Giftigkeit heranziehen kann. Um diese Zusammenhänge zu überprüfen, wurden verschiedene Kenngrößen ausgewählter Arten ermittelt (Tab. 2).
Tab. 2: verschiedene Kenngrößen ausgewählter Skorpionarten (Durchschnittswerte)2,3; der LD50-Wert gibt die Giftdosis an, bei der 50 % der Beutetiere sterben.
| Art | Körperlänge in mm | Scherendicke in mm | Schwanzdicke in mm | Dickenverhältnis | tödliche Giftdosis als LD50-Wert in mg/kg |
|---|---|---|---|---|---|
| Hottentotta juliae | 113 | 5,1 | 6,0 | 0,85 | 3,02 |
| Buthus awashensis | 54 | 1,9 | 4,7 | 0,40 | 0,90 |
| Heterometrus yaleensis | 75 | 11,6 | 4,4 | 2,64 | 300,00 |
| Pandinurus mazuchi | 93 | 10,5 | 4,5 | 2,33 | 40,00 |
| Parabuthus kabateki | 83 | 4,5 | 5,3 | 0,85 | 4,25 |
Beurteile unter Bezugnahme auf die Daten in Tabelle 2, ob die Körperlänge oder das Scheren-Schwanz-Dickenverhältnis eines Skorpions eine verlässlichere Aussage über die Giftigkeit der aufgeführten Skorpionarten zulässt.
Skorpione der Art Parabuthus villosus ernähren sich von Beutetieren, die in etwa ihrer Körpergröße entsprechen. Um Reptilien, Mäuse oder Insekten zu jagen, verlassen sie tagsüber ihre Verstecke und suchen sich einen geeigneten Platz, um der Beute aufzulauern. Mit Sinneshaaren nehmen diese Skorpione kleinste Erschütterungen am Boden in bis zu einem Meter Entfernung wahr und steuern dann zielgerichtet auf die Beute zu. Haben sie die Tiere mit den Scheren gepackt, wird die Beute sehr genau abgetastet. Passt das Tier ins Beutespektrum, injizieren sie mit mehreren Stichen Gift an einer geeigneten Stelle. Die Beute wird anschließend ins Versteck gebracht und dort verzehrt. Eine erneute Jagd findet erst nach mehreren Tagen wieder statt.
Erkläre das Verhalten der Skorpione beim Beutefang unter Verwendung ethologischer Fachbegriffe. Gehe davon aus, dass es sich um eine erbkoordinierte Verhaltensweise handelt.
Plane ein Experiment, mit dem untersucht werden soll, ob die Körperbedeckung (Hornschuppen, Fell oder Chitinpanzer) für die Giftstiche nach vorherigem Ergreifen eines Beutetiers eine Rolle spielt.
Während der Paarungszeit geben die Weibchen mancher Skorpionarten ein Sexualpheromon an den Boden ab, das artgleiche Männchen anlockt und zur Begattung motiviert.
Interpretiere das beschriebene Verhalten der Skorpione auf Grundlage des Sender-Empfänger-Modells.
Stelle eine Kosten-Nutzen-Analyse der beschriebenen Kommunikation durch das Pheromon im Vergleich zur Kommunikation durch akustische Signale auf.
Weiter lernen mit SchulLV-PLUS!
monatlich kündbarSchulLV-PLUS-Vorteile im ÜberblickDu hast bereits einen Account?Chemische Synapse im Nervensystem eines Beutetiers

Postsynaptisches Potenzial ohne Gift
Bei einer normalen Signalübertragung am Synaptischen Spalt kommt es nach der Ankunft eines Aktionspotenzials an das Endknöpfchen zuerst zu einem Calciumeinstrom in die Zelle und eine darauf folgende Freisetzung von in Vesikeln gespeicherten Transmittern in den synaptischen Spalt. Nach der Neurotransmitterbindung an die postsynaptischen rezeptorgesteuerten Natriumionen-Kanäle öffnen sich diese kurzzeitig. Dadurch strömen Natriumionen in die Zelle ein und das Membranpotenzial depolarisiert für eine kurze Zeit, wodurch das typische exzitatorische postsynaptische Potenzial (EPSP) entsteht. Gleichzeitig werden die Transmitter im synaptischen Spalt jedoch auch direkt wieder abgebaut bzw. zurück in das Endknöpfchen transportiert, was nach einer Zeit eine erneute Schließung der Kanäle bewirkt. Sobald dies geschieht, fließen die Membranströme der Zelle, insbesondere der Kaliumausstrom über Leckkanäle, und das Membranpotenzial kehrt rasch zum Ruhewert zurück. In der Abbildung zeigt sich deshalb ein kurzer, schmaler Ausschlag, der schnell wieder abfällt.
Postsynaptisches Potenzial mit Gift
Wird α-Toxin verabreicht, öffnen sich direkt die postsynaptischen Natriumionen-Kanäle (unabhängig ob davor schon ein Aktionspotenzial an das Endknöpfchen gelangt ist), da das Toxin daran bindet und die Kanäle daueröffnet. Der kontinuierliche Natriumeinstrom hält die postsynaptische Membran dauerhaft depolarisiert, sodass das Potenzial schnell steigt und sich dem Natriumgleichgewichtspotenzial annähert. Die Kurve fällt nicht wie im Normalfall schnell zum Ruhepotenzial zurück, sondern verharrt auf einem stark depolarisierten Niveau und sinkt nur langsam, da es keinen schnellen Response- (Abbau- und Abtransport-) mechanismus gibt, wie bei dem Neurotransmitter im Normalfall.

Aktionspotenzial und Ioentheorie im Normalfall
Im unveränderten Axonverlauf entsteht ein Aktionspotenzial, weil sich spannungsabhängige Natriumionenkanäle öffnen. Dies geschieht nach dem „alles oder nichts Prinzip“: Die Membran des entsprechenden Axons muss von eingehenden Signalen über einen gewissen Schwellenwert hinaus depolarisiert werden, dann leitet sich das Signal als Aktionspotenzial weiter. Das Ruhepotenzial, also die elektrische Spannung zwischen der Innen- und Außenseite einer unerregten Zelle, das laut Ionentheorie durch die ungleiche Verteilung von Ionen und somit Ladungen auf beiden Seiten der semipermeablen Membran entsteht, wird somit aufgehoben. Natriumionen strömen nun also bei Start eines Aktionspotenzials entlang ihres elektrochemischen Gradienten in das Axon ein und depolarisieren die Membran bis in den positiven Bereich. Unmittelbar danach öffnen sich spannungsabhängige Kaliumionenkanäle. Kaliumionen verlassen die Zelle entlang ihres Gradienten, wodurch das Membranpotenzial wieder in den negativen Bereich zurückkehrt. Dieser Kaliumausstrom sorgt für die Repolarisation und meist eine kurze Hyperpolarisation. Erst wenn das Ruhepotenzial wieder annähernd erreicht ist, können weitere Aktionspotenziale in normaler Weise ausgelöst und entlang des Axons weitergeleitet werden.
Wirkung von Agitoxin 1
Agitoxin 1 hemmt die Kaliumionenkanäle in der Axonmembran und blockiert damit den für die Repolarisation notwendigen Kaliumausstrom. Nach der Depolarisation durch den Natriumeinstrom bleibt das Membranpotenzial länger auf einem deutlich positiveren Niveau, weil der Ausgleich über Kaliumionen ausbleibt oder stark verzögert ist. Die Folge ist eine verlängerte Depolarisationsphase und eine erschwerte Rückkehr zum Ruhepotenzial. Dadurch bleiben viele spannungsabhängige Natriumkanäle in einem inaktiven Zustand und das Axon kann nur eingeschränkt oder gar keine neuen Aktionspotenziale bilden. Die Erregungsleitung wird instabil, bis hin zum völligen Leitungsblock, was beim Beutetier zu Lähmungserscheinungen und letztlich zum Ausfall lebenswichtiger Funktionen führen kann.
Vergleich der Faustregeln zur Giftigkeit von Skorpionen
Der LD50-Wert gibt die Giftdosis in mg pro kg Körpergewicht an, bei der 50 Prozent der Versuchstiere sterben. Je niedriger der LD50-Wert ist, desto weniger Gift wird bis zum Tod benötigt und desto gefährlicher beziehungsweise giftiger ist das Toxin für die Beute. Ein Skorpion mit einem LD50 von 0,90 mg/kg (vgl. Buthus awashensis) besitzt also ein deutlich stärkeres Gift als ein Skorpion mit einem LD50 von 40 mg/kg (vgl. Pandinurus mazuchi).
Betrachtet man nun die Faustregel zur Körperlänge, nach der kleinere Skorpione giftiger sein sollen, lassen sich aus den Daten nur teilweise passende Zusammenhänge ableiten. Buthus awashensis ist mit 54 mm Körperlänge der kleinste der aufgeführten Skorpione und hat mit 0,90 mg/kg tatsächlich den niedrigsten LD50-Wert. Hottentotta juliae ist mit 113 mm der größte Skorpion der Tabelle und weist mit 3,02 mg/kg ebenfalls ein recht starkes Gift auf. Heterometrus yaleensis ist 75 mm lang und damit kleiner als Hottentotta juliae, nun zeigt sich die postulierte Regel jedoch als fehlbar, da die kleinere Art Heterometrus yaleensis mit einem LD50-Wert von 300 mg/kg um ein Vielfaches ungiftiger ist. Es gibt daher kleine, sehr giftige Arten, aber auch größere Arten mit starkem Gift und mittelgroße Arten mit sehr schwachem Gift. Die Körperlänge allein erlaubt somit keine zuverlässige Vorhersage der Giftigkeit.
Deutlicher ist der Zusammenhang beim Scheren-Schwanz-Dickenverhältnis. Heterometrus yaleensis besitzt das höchste Dickenverhältnis mit 2,64 und ist mit 300 mg/kg am wenigsten giftig. Buthus awashensis besitzt das niedrigste Dickenverhältnis von 0,40 und ist mit 0,90 mg/kg am giftigsten. Hottentotta juliae und Parabuthus kabateki haben mittlere, gleiche Dickenverhältnisse von jeweils 0,85 und zeigen mit 3,02 mg/kg beziehungsweise 4,25 mg/kg ebenfalls eine hohe Giftigkeit. Daraus ergibt sich als Faustregel: Je kleiner das Scheren-Schwanz-Dickenverhältnis ist, desto stärker ist das Gift.
Dieses Dickenverhältnis erlaubt daher eine deutlich verlässlichere Einschätzung der Giftigkeit als die reine Körperlänge.
Skorpionverhalten bei Beutefang
Das beschriebene Beutefangverhalten der Skorpione lässt sich als erbkoordinierte Verhaltensweise einordnen, was bedeutet dass es artspezifisch, angeboren und ohne vorheriges Lernen vollständig ausgeführt wird. Die Jagd beginnt mit einem angeborenen Auslösemechanismus (AAM), der durch bestimmte Schlüsselreize aktiviert wird. Solche Schlüsselreize sind in diesem Fall feinste Bodenerschütterungen, die die Skorpione mit ihren Sinneshaaren wahrnehmen. Diese Reize lösen ein appetitives Verhalten aus, das sich durch Suchen, Orientierung und Annäherung an die potenzielle Beute zeigt. Die zielgerichtete Bewegung auf den Ursprung der Erschütterung zu ist Teil dieser Such- und Orientierungsphase.
Hat der Skorpion die Beute ergriffen, geht das Verhalten in eine endhandlungstypische Phase über, die durch ein festgelegtes Bewegungsmuster gekennzeichnet ist. Das Abtasten der Beute dient als weiterer Schlüsselreiz, der überprüft, ob das Tier in das artspezifische Beutespektrum fällt. Wird dieser innere Filter positiv erfüllt, wird die Endhandlung, nämlich das Injizieren des Gifts durch mehrere Stiche, ausgelöst. Diese Endhandlung ist ungestört und stereotyp, typisch für eine erbkoordinierte Bewegung. Anschließend folgt ein ebenfalls festgelegtes Verhalten: das Transportieren der Beute ins Versteck und eine mehrtägige Jagdpause, was den Handlungsablauf einer Instinkthandlung komplettiert.
Fragestellung
Beeinflusst die Art der Körperbedeckung eines Beutetiers die Durchführung der Giftstiche durch Parabuthus villosus nach dem Ergreifen der Beute? Um dies zu prüfen, wird ein kontrolliertes verhaltensbiologisches Experiment durchgeführt.
Planung des Experiments
Dazu verwendet man drei künstliche oder tote Beutekörper gleicher Größe und Form, die jeweils eine typische Körperbedeckung nachbilden: Hornschuppen zur Simulation eines Reptils, Fell zur Simulation eines Säugetiers und einen Chitinpanzer zur Simulation eines Insekts. Alle anderen Eigenschaften wie Gewicht, Temperatur, Materialhärte und die Art des mechanischen Reizes beim Ergreifen werden konstant gehalten, um sicherzustellen, dass nur die Körperbedeckung als unabhängige Variable variiert.
Jeder Skorpion erhält nur ein Beutemodell, das ihm im Terrarium präsentiert wird. Sobald der Skorpion das Objekt mit seinen Scheren ergriffen hat, beginnt die Beobachtung. Innerhalb eines festen Beobachtungszeitraums wird dokumentiert, wie viele Giftstiche ausgeführt werden, wie lange der Skorpion bis zur Stichabgabe benötigt und an welchen Stellen des Beutekörpers die Stiche gesetzt werden. Die Versuche werden mit einer ausreichend großen Zahl an Individuen wiederholt, um zufällige Effekte auszuschließen.
In der anschließenden Auswertung werden die Reaktionen der Skorpione zwischen den drei Gruppen verglichen. Zeigen sich deutliche Unterschiede in der Anzahl oder Position der Stiche, kann daraus gefolgert werden, dass die Art der Körperbedeckung die Endhandlung des Beiß- und Stechverhaltens beeinflusst. Bleiben Unterschiede aus, spricht dies dafür, dass die Körperbedeckung keine wesentliche Rolle bei der Durchführung der Giftstiche spielt.
Paarungsverhalten nach Sender-Empfänger-Modell
Während der Paarungszeit fungieren die Weibchen als Sender im Sinne des Sender-Empfänger-Modells. Sie geben ein artspezifisches Sexualpheromon an den Boden ab und kodieren damit ihre Paarungsbereitschaft und ihre Anwesenheit. Dieses Pheromon stellt das chemische Signal dar, das über das Medium Boden und Luft verbreitet wird und so einen chemischen Kommunikationskanal bildet. Die Konzentration und Zusammensetzung des Pheromons sorgt dafür, dass in erster Linie artgleiche Männchen angesprochen werden, was die Spezifität der Kommunikation erhöht.
Die Männchen übernehmen die Rolle der Empfänger. Sie besitzen spezialisierte Chemorezeptoren, mit denen sie das Pheromon wahrnehmen und die chemische Information decodieren. Wird eine bestimmte Reizschwelle überschritten, löst dies beim Männchen ein gerichtetes Such- und Orientierungsverhalten hin zum Weibchen aus, das in der Paarung als Reaktion auf das empfangene Signal mündet. Damit liegt eine erfolgreiche Kommunikation im Sinne des Sender-Empfänger-Modells vor, bei der der Sender durch das Pheromon ein Signal aussendet, der Empfänger dieses Signal über einen chemischen Kanal aufnimmt, verarbeitet und sein Verhalten zielgerichtet anpasst.
Kosten-Nutzen-Analyse der beschriebenen Kommunikation
Eine chemische Kommunikation über Pheromone hat für die Weibchen zunächst einen wesentlichen Nutzen, da das ausgesandte Sexualpheromon sehr artspezifisch wirkt und nur die passenden Männchen anzieht. Zudem ist die chemische Kommunikation energetisch kostengünstig, da die Produktion kleiner Pheromonmengen weniger Energie verbraucht als das Erzeugen lauter akustischer Signale. Ein weiterer Vorteil besteht darin, dass Pheromone über längere Zeit im Substrat bestehen bleiben, sodass auch später vorbeikommende Männchen noch erreicht werden können. Damit erhöht sich die Wahrscheinlichkeit einer erfolgreichen Paarung deutlich. Demgegenüber stehen Kosten, da ein Pheromon nicht nur geeignete Männchen, sondern unter Umständen auch Räuber oder konkurrierende Skorpione anlocken kann. Außerdem können Pheromonsignale bei ungünstigen Umweltbedingungen, etwa starkem Wind oder sehr trockenen Böden, weniger effektiv verbreitet oder schneller abgebaut werden.
Im Vergleich dazu hat eine akustische Kommunikation ebenfalls Vorteile: Sie kann eine größere Reichweite besitzen und erreicht Männchen schneller. Allerdings sind akustische Signale wesentlich energieaufwändiger herzustellen und machen den Sender leicht lokalisierbar, was das Risiko erhöht, selbst zur Beute zu werden. Zudem ist die Spezifität akustischer Signale oft geringer als die chemischer Pheromone, was zu Fehlkommunikation führen kann.
Insgesamt überwiegt unter den gegebenen Bedingungen der Nutzen der chemischen Pheromonsignale, da sie spezifischer, sparsamer und in der Paarungssituation der Skorpione effektiver sind als akustische Signale.