C2 Moore
Hinweis: In der Prüfung musst du von den Aufgabenblöcken A, B und C jeweils einen der beiden Vorschlägen 1 oder 2 bearbeiten.
Moore sind Feuchtgebiete, in denen sich durch die Stoffwechselvorgänge von Torfmoosen (Sphagnum) und unterschiedlichen Bakterien organisches Material ansammelt. Dabei wächst das Moor durch immer neue Torfmoosschichten stetig an.
Die Fotosyntheseaktivität jeder Moospflanze hängt stark vom jeweiligen Standort innerhalb des Moores ab.
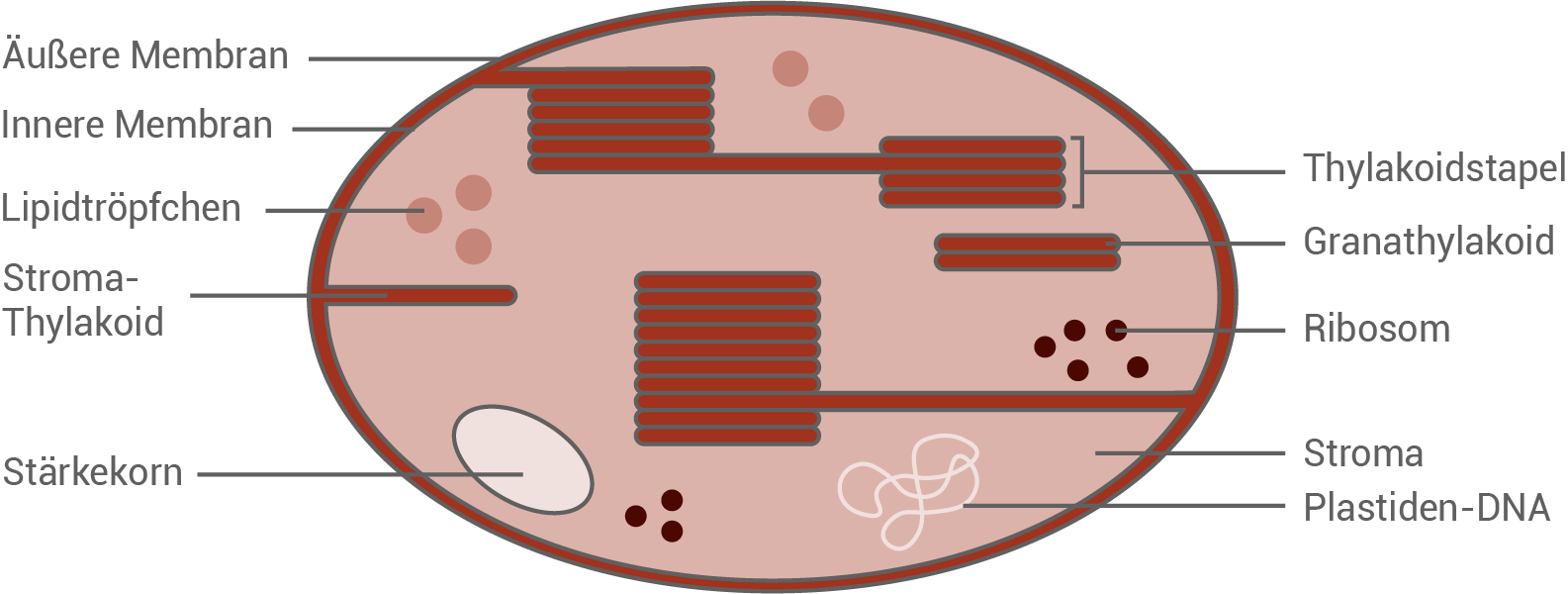
Fertige eine beschriftete Skizze des elektronenmikroskopischen Aufbaus des fotosynthetisch aktiven Zellorganells einer Torfmoospflanze an.
In einer Studie untersuchten Forschende die fotosynthetischen Reaktionen von Sphagnum-Pflanzen bei unterschiedlicher Wasserverfügbarkeit. Bei hoher Wasserverfügbarkeit erkennt man, dass die Pflanze mit einem Wasserfilm überzogen ist, der die Kohlenstoffdioxid-Verfügbarkeit in der Pflanze herabsetzt. Forschende haben diesen Einfluss des Wasserfilms mit einem Experiment überprüft.
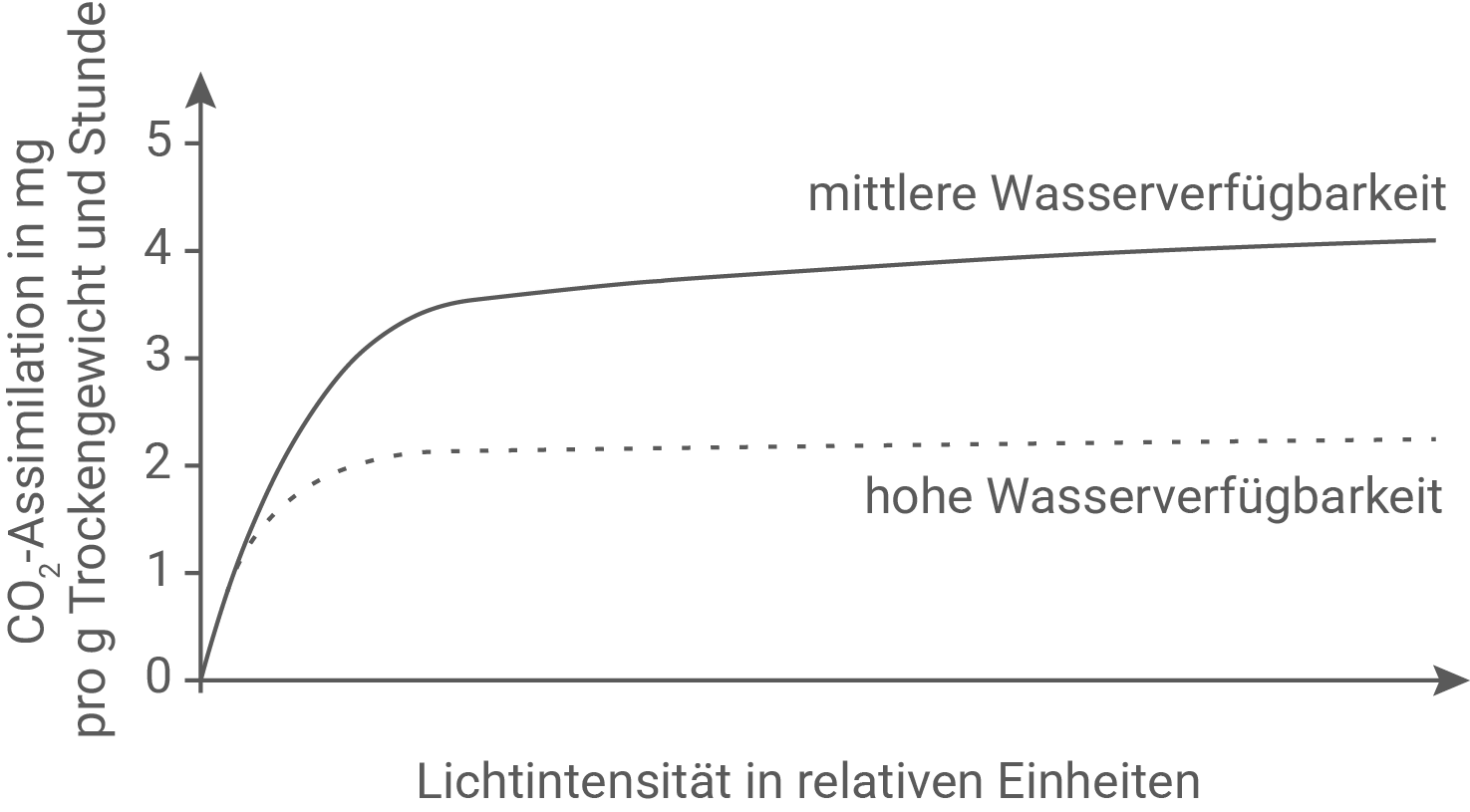
Beschreibe eine mögliche Durchführung eines Experiments, durch das die in Abbildung 1 gezeigten Ergebnisse ermittelt werden könnten. Nenne die fotosynthetischen Reaktionen, die durch die reduzierte Kohlenstoffdioxid-Verfügbarkeit beeinträchtigt werden und gib die dazugehörige Bruttogleichung an.
Der Großteil der Sphagnum-Pflanzen steht im Moor stets unter Wasser. Dadurch ergeben sich mehrere Schichten im Moor mit jeweils typischen Stoffwechselprozessen. So kommt es neben der Fotosynthese von Moospflanzen auch zu Stoffwechselprozessen von Bakterien (Abb. 2).
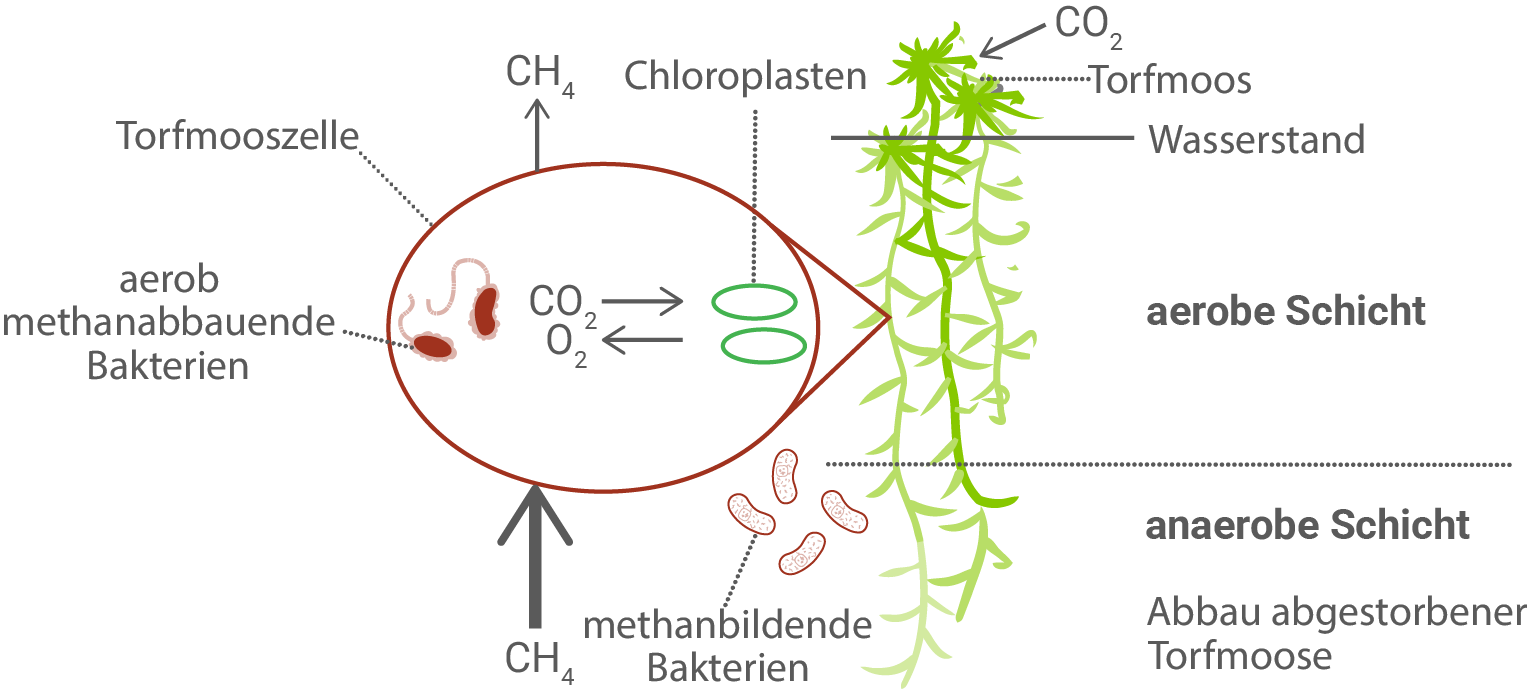
Erläutere jeweils einen Vorteil für Sphagnum-Arten und methanabbauende Bakterien durch das in Abbildung 2 dargestellte Zusammenleben.
Torf bildet sich über lange Zeiträume durch den langsamen enzymatischen Abbau von Pflanzenmaterial und besteht unter anderem aus Cellobiose. Als Nebenprodukt dieses Abbaus können Polyphenole entstehen. Während in gut durchlüfteter Erde der Abbau von Cellobiose durch das Enzym β-Glucosidase schnell verläuft (Abb. 3), wird er bei der Torfbildung in den anaeroben Schichten des Moores durch das Vorkommen von Polyphenolen gehemmt. Bei Anwesenheit von Sauerstoff werden Polyphenole durch das Enzym Phenoloxidase abgebaut (Abb. 3).
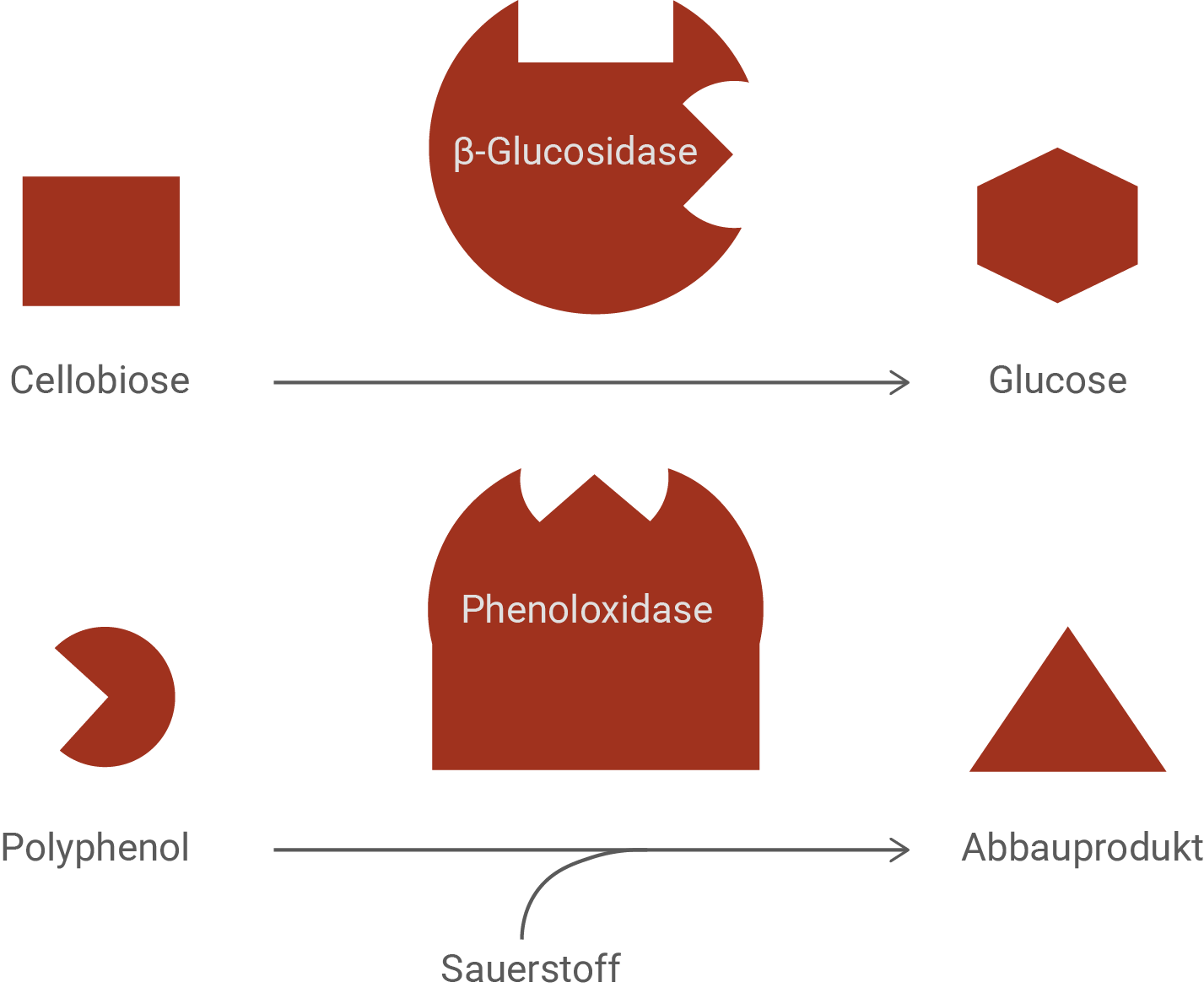
Erläutere auf Teilchenebene die Art der bei der Torfbildung vorliegenden Hemmung. Gehe dabei auf die Notwendigkeit anaerober Bedingungen ein.
Durch die Assimilation von Kohlenstoffdioxid sind Ökosysteme in unterschiedlichem Maße in der Lage, Kohlenstoff zu speichern und damit die Menge von klimaschädlichen Gasen in der Atmosphäre zu reduzieren.
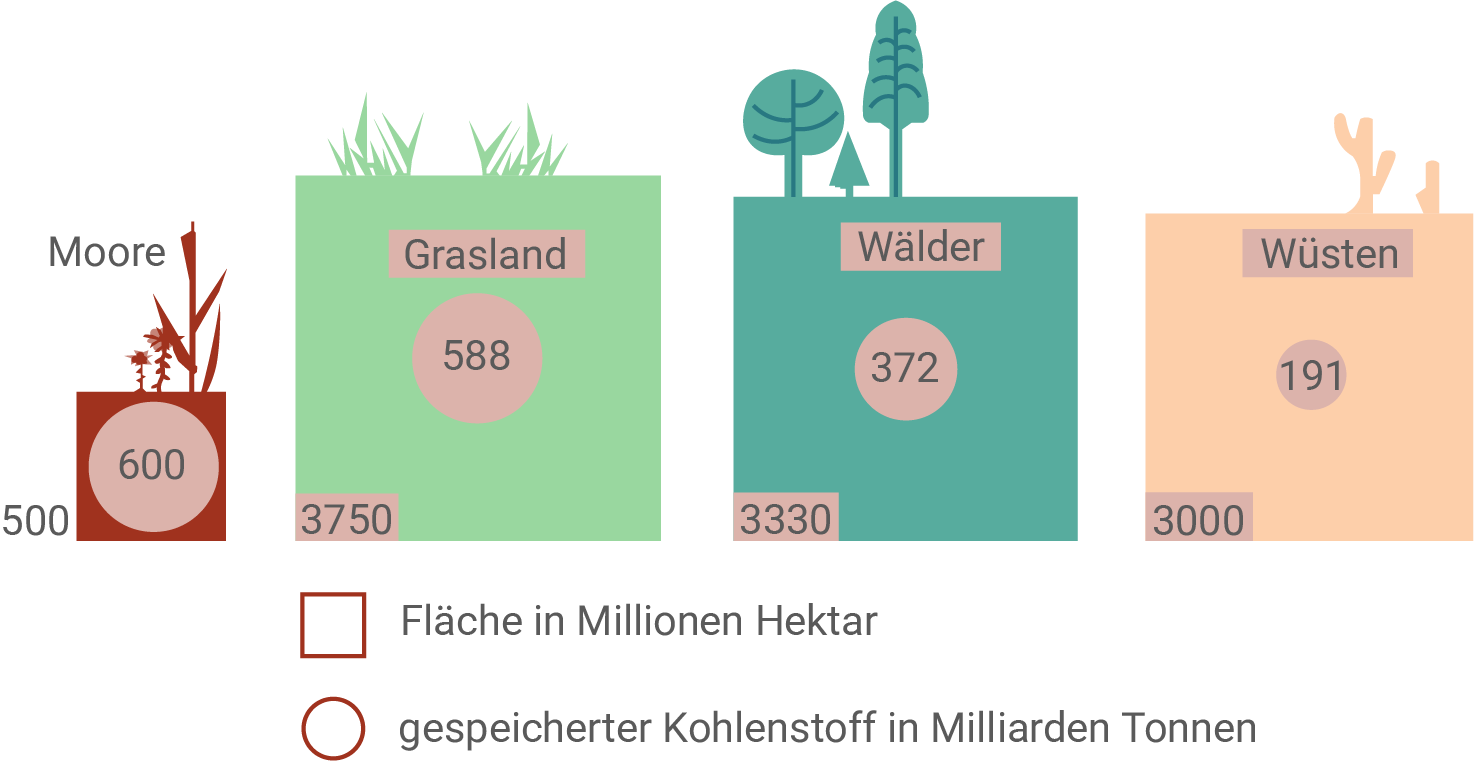
Erläutere unter Zuhilfenahme von Abb. 4 die Auswirkungen einer Umwandlung von Grasland zu Mooren auf die Kohlenstoffdioxidbilanz.
Das Moor ist u. a. für Insekten und Vögel ein vielfältiger Lebensraum. Auch um diese Lebensräume zu schützen, werden vormals trocken gelegte Moore wieder vernässt. In solchen renaturierten Feuchtgebieten siedelt sich die Gemeine Stechmücke (Culex pipiens) an. Deren Weibchen legen jeweils etwa 150-300 Eier. Vom Ei bis zur geschlechtsreifen Stechmücke dauert die Entwicklung 10-14 Tage. In nur kurzer Zeit etabliert sich somit eine stabile Population. Auch Wirbeltiere wie die Moorente (Aythya nyroca) siedeln sich an. Sie legt 7-11 Eier, die vom Weibchen 3-4 Wochen ausgebrütet werden. Die geschlüpften Jungtiere sind Nestflüchter, werden jedoch noch etwa zwei Monate durch die Mutter beschützt. Mit einem Alter von einem Jahr werden die Tiere geschlechtsreif.
Erstelle ein Diagramm, das die Entwicklung einer Mückenpopulation in einem kürzlich wieder vernässten Moor aufzeigt, und ordne diesem die einzelnen Phasen der Populationsentwicklung zu.
Erkläre zwei Unterschiede der Populationsentwicklung einer Mücken- und einer Moorentenpopulation unter Berücksichtigung der Fortpflanzungsstrategien beider Tierarten.
Weiter lernen mit SchulLV-PLUS!
monatlich kündbarSchulLV-PLUS-Vorteile im ÜberblickDu hast bereits einen Account?Elektronenmikroskopischer Aufbau eines Chromoplasten
Experiment zur Reproduktion der Messdaten
Eine mögliche experimentelle Durchführung, mit der die in Abbildung 1 dargestellten Kurven ermittelt werden können, besteht darin, gleich große und möglichst ähnliche Sphagnum-Proben unter standardisierten Bedingungen in eine Messkammer zu bringen und die Netto-CO2-Assimilation bei variierender Lichtintensität zu bestimmen. Dazu werden zunächst mehrere Proben mit gleicher Frischmasse oder gleicher Oberfläche vorbereitet und anschließend auf definierte Wasserverfügbarkeitsstufen eingestellt. Für die Bedingung „mittlere Wasserverfügbarkeit“ werden die Proben so befeuchtet, dass sie hydratisiert sind, aber kein zusammenhängender Wasserfilm die Oberfläche dauerhaft bedeckt. Für die Bedingung „hohe Wasserverfügbarkeit“ werden die Proben so stark befeuchtet, dass ein sichtbarer Wasserfilm auf der Pflanzenoberfläche entsteht. Damit die Wasserstufen während der Messung konstant bleiben, kann man die Proben auf inertem Substrat platzieren und die Luftfeuchtigkeit sowie die Temperatur in der Messkammer konstant halten, sodass Verdunstung kontrollierbar ist. Anschließend wird die Kammer mit Luft definierter CO2-Konzentration und konstanter Temperatur durchströmt. Die CO2-Assimilation wird über die Differenz der CO2-Konzentration zwischen einströmender und ausströmender Luft gemessen, beispielsweise mit einem Infrarot-Gasanalysator. Parallel wird die Lichtintensität stufenweise erhöht, etwa von sehr geringer Beleuchtung bis zur Sättigungsbeleuchtung. Für jede Lichtstufe wird nach einer kurzen Anpassungszeit die stabile CO2-Aufnahme pro Zeit bestimmt und auf Trockenmasse bezogen, wie es die y-Achse der Abbildung zeigt. So erhält man für jede Wasserbedingung eine Lichtantwortkurve. Erwartungsgemäß steigt die CO2-Assimilation zunächst mit zunehmender Lichtintensität an und erreicht dann ein Plateau. Bei hoher Wasserverfügbarkeit liegt dieses Plateau deutlich niedriger als bei mittlerer Wasserverfügbarkeit, was damit vereinbar ist, dass ein Wasserfilm den CO2-Zutritt in das Pflanzengewebe verringert und damit das CO2-Angebot im Inneren der Pflanze limitiert.
Beeinträchtigte fotosynthetischen Reaktionen
Die reduzierte Kohlenstoffdioxid-Verfügbarkeit beeinträchtigt nicht die lichtabhängigen Reaktionen direkt, sondern vor allem die lichtunabhängigen Reaktionen der Fotosynthese, also den Calvin-Zyklus im Stroma der Chloroplasten. Dort wird CO2 durch das Enzym Rubisco an Ribulose-1,5-bisphosphat gebunden, und die entstehenden Zwischenprodukte werden unter Verbrauch von ATP und NADPH zu Triosephosphaten reduziert, aus denen Kohlenhydrate entstehen. Wenn weniger CO2 verfügbar ist, sinkt die Carboxylierungsrate der Rubisco, und dadurch wird die gesamte CO2-Fixierung begrenzt. In der Folge werden ATP und NADPH langsamer verbraucht, NADP+ wird langsamer regeneriert, und damit kann auch die lichtabhängige Elektronentransportkette indirekt „gebremst“ werden, weil weniger NADP+ als terminaler Elektronenakzeptor zur Verfügung steht. Zusätzlich begünstigt ein niedriger CO2-Gehalt im Verhältnis zum O2-Gehalt die Oxygenase-Reaktion der Rubisco, wodurch Photorespiration zunimmt. Photorespiration führt dazu, dass bereits gebundener Kohlenstoff teilweise wieder als CO2 freigesetzt wird und zusätzlich ATP verbraucht wird, ohne dass Zucker aufgebaut wird. Das senkt die Netto-Assimilation weiter, besonders unter Bedingungen, in denen CO2 schlecht in das Gewebe diffundiert.
Die Bruttogleichung der lichtunabhängigen Reaktion der Fotosynthese lautet:
Vorteile durch das dargestellte Zusammenleben
In Abbildung 2 ist ein Moor mit einer aeroben Schicht oberhalb des Wasserstands und einer anaeroben Schicht darunter dargestellt. In der anaeroben Schicht bauen methanbildende Bakterien abgestorbenes Torfmoos unter Sauerstoffausschluss ab und setzen dabei Methan frei, das nach oben diffundiert. In der aeroben Schicht leben methanabbauende Bakterien, die dieses Methan mit Sauerstoff zu Kohlenstoffdioxid oxidieren. Das dabei entstehende Kohlenstoffdioxid steht wiederum dem Torfmoos als Substrat für die Fotosynthese zur Verfügung.
Für Sphagnum-Arten ergibt sich aus dieser Beziehung der Vorteil, dass sie zusätzliches Kohlenstoffdioxid erhalten. Da im Moor insbesondere bei hoher Wasserverfügbarkeit die CO2-Diffusion aus der Luft in das Pflanzengewebe erschwert sein kann, wirkt das von methanabbauenden Bakterien gebildete CO2 wie eine lokale CO2-Quelle in unmittelbarer Umgebung der Torfmoospflanzen. Dadurch kann die CO2-Assimilation in den Chloroplasten unterstützt werden, was den Aufbau von Kohlenhydraten und damit das Wachstum begünstigt.
Für die methanabbauenden Bakterien liegt der Vorteil darin, dass sie durch das Torfmoos geeignete Bedingungen und Substrate erhalten. Die Pflanze liefert in der aeroben Zone Sauerstoff, der durch die Fotosynthese entsteht und für die Methanoxidation benötigt wird. Das Zusammenleben ist damit für die Bakterien energetisch vorteilhaft, weil sie aus der Oxidation von Methan Energie gewinnen können und gleichzeitig über das durch die Pflanze bereitgestellte sauerstoffreiche Milieu in der oberen Schicht optimale Voraussetzungen für diesen Stoffwechsel haben.
Enzymhemmung bei Torfbildung
Aus der Aufgabenstellung ist zu erkennen, dass der Abbau von Cellobiose zu Glucose durch das Enzym β-Glucosidase grundsätzlich möglich ist, in den anaeroben Moorschichten jedoch durch das Auftreten von Polyphenolen stark verlangsamt wird. Auf Teilchenebene lässt sich diese Hemmung so erklären, dass Polyphenole als Hemmstoff mit dem Enzym in Wechselwirkung treten und dadurch die Umsetzung des Substrats verhindern. Aus der gegebenen Abbildung lässt sich herleiten, dass es sich dabei nicht um eine klassische kompetitive Hemmung handelt, bei der der Hemmstoff das aktive Zentrum direkt blockiert, sondern um eine Hemmung durch Bindung an einer anderen Stelle des Enzyms, die dessen räumliche Struktur und damit die Funktion des aktiven Zentrums verändert. Polyphenole können sich an das Enzym anlagern (vergleiche passendes „Loch“ als allosterische Bindungsstelle an der β-Glucosidase) und über Konformationsänderungen nach Bindung dafür sorgen, dass Cellobiose entweder schlechter binden kann oder dass der Enzym-Substrat-Komplex nicht mehr effizient zu Produkt umgesetzt wird.
Die Notwendigkeit anaerober Bedingungen ergibt sich aus dem zweiten Teil der Abbildung. Polyphenole werden in Anwesenheit von Sauerstoff durch das Enzym Phenoloxidase abgebaut. Sauerstoff wirkt dabei als notwendiger Reaktionspartner, sodass Phenoloxidase unter aeroben Bedingungen Polyphenole zu Abbauprodukten umsetzen kann. In gut durchlüfteter Erde werden Polyphenole daher entfernt, die Hemmwirkung nimmt ab und β-Glucosidase kann Cellobiose wieder schnell zu Glucose spalten. In den anaeroben Schichten des Moores fehlt Sauerstoff, wodurch die Phenoloxidase nicht oder nur stark eingeschränkt arbeiten kann. Polyphenole bleiben deshalb erhalten, reichern sich an und hemmen dauerhaft den enzymatischen Abbau pflanzlichen Materials. Genau diese Kombination aus Sauerstoffmangel und Hemmstoffpersistenz führt dazu, dass der Abbau organischer Substanz sehr langsam abläuft und sich über lange Zeiträume Torf ansammeln kann.
Auswirkungen einer Umwandlung von Grasland zu Mooren
Abbildung 4 zeigt, dass Moore im Verhältnis zu ihrer globalen Fläche besonders viel Kohlenstoff speichern. Während Grasland weltweit eine Fläche von etwa 3750 Millionen Hektar einnimmt und insgesamt ungefähr 588 Milliarden Tonnen Kohlenstoff speichert, umfassen Moore nur rund 500 Millionen Hektar, speichern aber etwa 600 Milliarden Tonnen Kohlenstoff. Damit liegt die gespeicherte Kohlenstoffmenge der Moore trotz deutlich geringerer Fläche sogar höher als die des Graslands. Das bedeutet, dass Moore bezogen auf die Fläche eine deutlich größere Kohlenstoffdichte besitzen und langfristig wesentlich mehr Kohlenstoff in Form von Torf binden können.
Wird Grasland in Moor umgewandelt, führt dies daher langfristig zu einer stärkeren Entnahme von Kohlenstoffdioxid aus der Atmosphäre und zu einer verbesserten Kohlenstoffdioxidbilanz, weil in Mooren unter den anaeroben Bedingungen der Zersetzungsprozess stark gehemmt ist und organisches Material über lange Zeit als Torf akkumuliert. In der Bilanz bedeutet das, dass ein größerer Anteil des durch Fotosynthese aufgenommenen Kohlenstoffs nicht wieder als CO2 freigesetzt wird, sondern als organischer Kohlenstoff im Boden gespeichert bleibt. Die Zahlen aus der Abbildung stützen diese Aussage, denn obwohl Moore nur etwa ein Siebtel bis ein Achtel der Graslandfläche besitzen, speichern sie insgesamt ähnlich viel beziehungsweise sogar mehr Kohlenstoff. Eine Umwandlung würde somit die Speicherung pro Fläche erhöhen und die atmosphärische CO2-Konzentration tendenziell senken, weil mehr Kohlenstoff dauerhaft aus dem kurzfristigen Kohlenstoffkreislauf in einen langfristigen Speicher überführt wird.
Hinweis: Zur Bearbeitung der Aufgabe kann auch mit den ausgerechneten Werten an Kohlenstoff pro Flächeneinheit der jeweiligen Ökosysteme argumentiert und diese untereinander verglichen werden.
Diagramm zur Entwicklung der Mückenpopulation

Hinweis: Der Graph stellt die Populationsentwicklung qualitativ dar. Die einzelnen dargestellten Phasen können in der Realität in ihrer Länge und Ausgeprägtheit abweichen.
Vergleich der Populationsentwicklung
Ein erster Unterschied betrifft die Geschwindigkeit des Populationswachstums. Die Stechmücke folgt einer r-Strategie: Sie legt sehr viele Eier, zwischen etwa 150 und 300 pro Weibchen, und die Entwicklung vom Ei bis zum geschlechtsreifen Tier dauert nur 10 bis 14 Tage. Dadurch können sich in kurzer Zeit viele Individuen fortpflanzen, sodass die Population unter günstigen Umweltbedingungen sehr schnell anwächst und sich rasch etabliert. Die Moorente hingegen ist ein typischer K-Strategist. Sie legt deutlich weniger Eier, meist nur 7 bis 11, und die Entwicklung ist langwierig, da die Brutzeit mehrere Wochen dauert und die Jungtiere noch über Monate betreut werden. Zudem erreichen die Tiere die Geschlechtsreife erst nach etwa einem Jahr. Dadurch wächst die Population der Moorente wesentlich langsamer und reagiert verzögert auf günstige Umweltbedingungen.
Ein zweiter Unterschied liegt in der Stabilität und Regulation der Population. Bei der Stechmücke ist die individuelle Überlebenswahrscheinlichkeit gering, da keine Brutpflege erfolgt und viele Larven oder Jungtiere früh sterben. Die Populationsgröße schwankt daher stark und ist stark von Umweltfaktoren wie Wasserverfügbarkeit oder Prädation abhängig. Bei der Moorente ist die Überlebenswahrscheinlichkeit der einzelnen Nachkommen deutlich höher, da die Eier bebrütet und die Jungtiere über längere Zeit geschützt werden. Die Population ist deshalb insgesamt stabiler, schwankt weniger stark und nähert sich langsamer der Umweltkapazität an.
Zusammengefasst wächst die Mückenpopulation aufgrund hoher Nachkommenzahl und kurzer Generationszeit schnell, aber instabil, während die Moorentenpopulation aufgrund geringer Nachkommenzahl, intensiver Brutpflege und langer Entwicklungszeit langsam, dafür aber vergleichsweise stabil zunimmt.