B1 Plankton
Hinweis: In der Prüfung musst du von den Aufgabenblöcken A, B und C jeweils einen der beiden Vorschlägen 1 oder 2 bearbeiten.
Zum Plankton zählt man alle lebenden Organismen, die im Wasser treiben und dabei von der Wasserströmung transportiert werden. Man unterteilt diese Gruppe von Lebewesen unter anderem in Zooplankton (Tiere), Phytoplankton (Pflanzen und Cyanobakterien) und Bakterioplankton (Bakterien).
Das Populationswachstum von Phytoplankton, wie das der Kieselalge Skeletonema marinoi, ist unter anderem abhängig von der Meersalzkonzentration. Um diesen Zusammenhang genauer zu untersuchen, wurden Proben von S. marinoi an zwei Stellen (BS, BA) mit jeweils unterschiedlicher Meersalzkonzentration entnommen (Abb. 1) und im Labor bei unterschiedlichen Salzkonzentrationen kultiviert. Die Ergebnisse dieser Untersuchungen zum Populationswachstum sind in Abbildung 2 dargestellt.
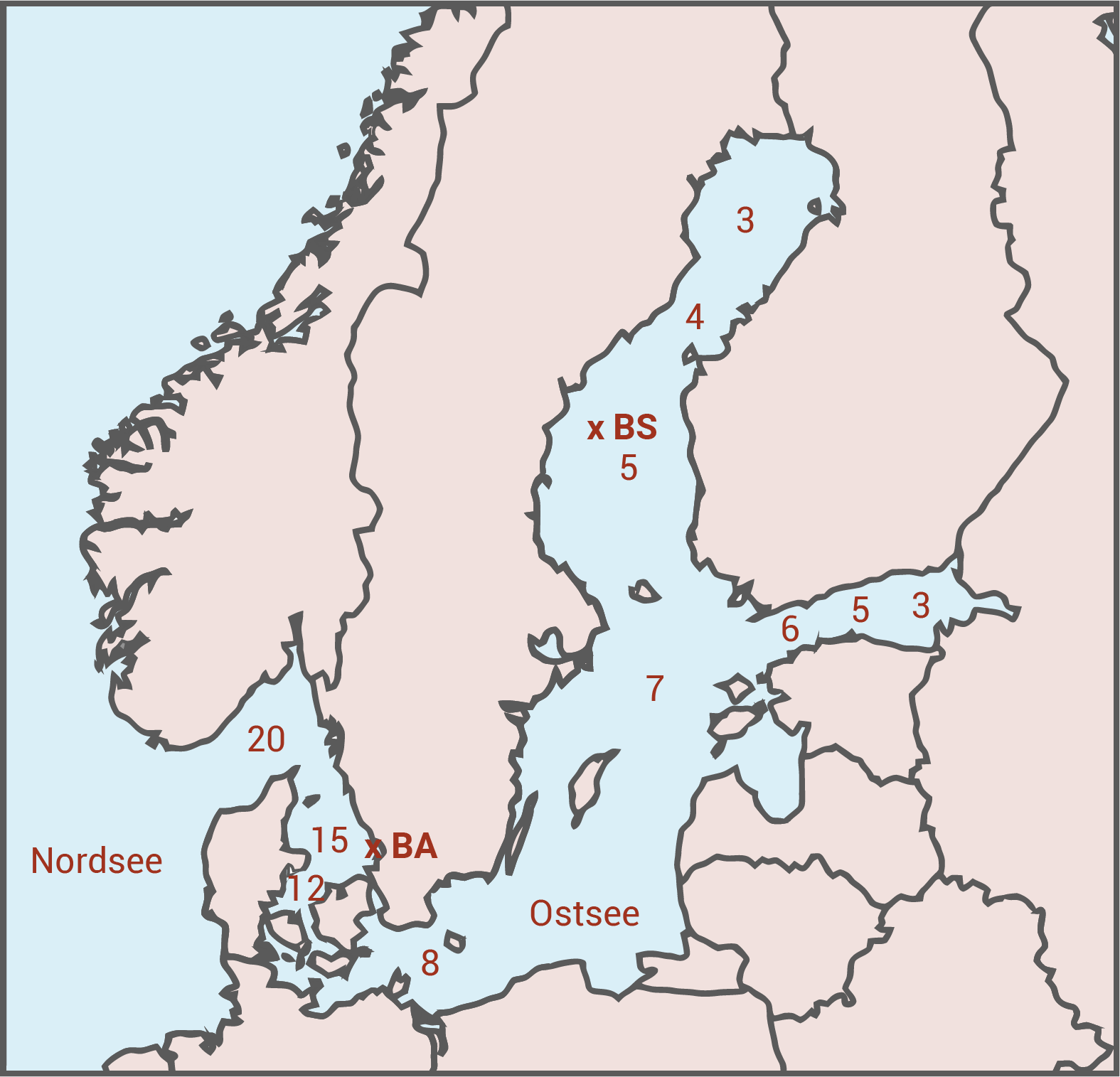
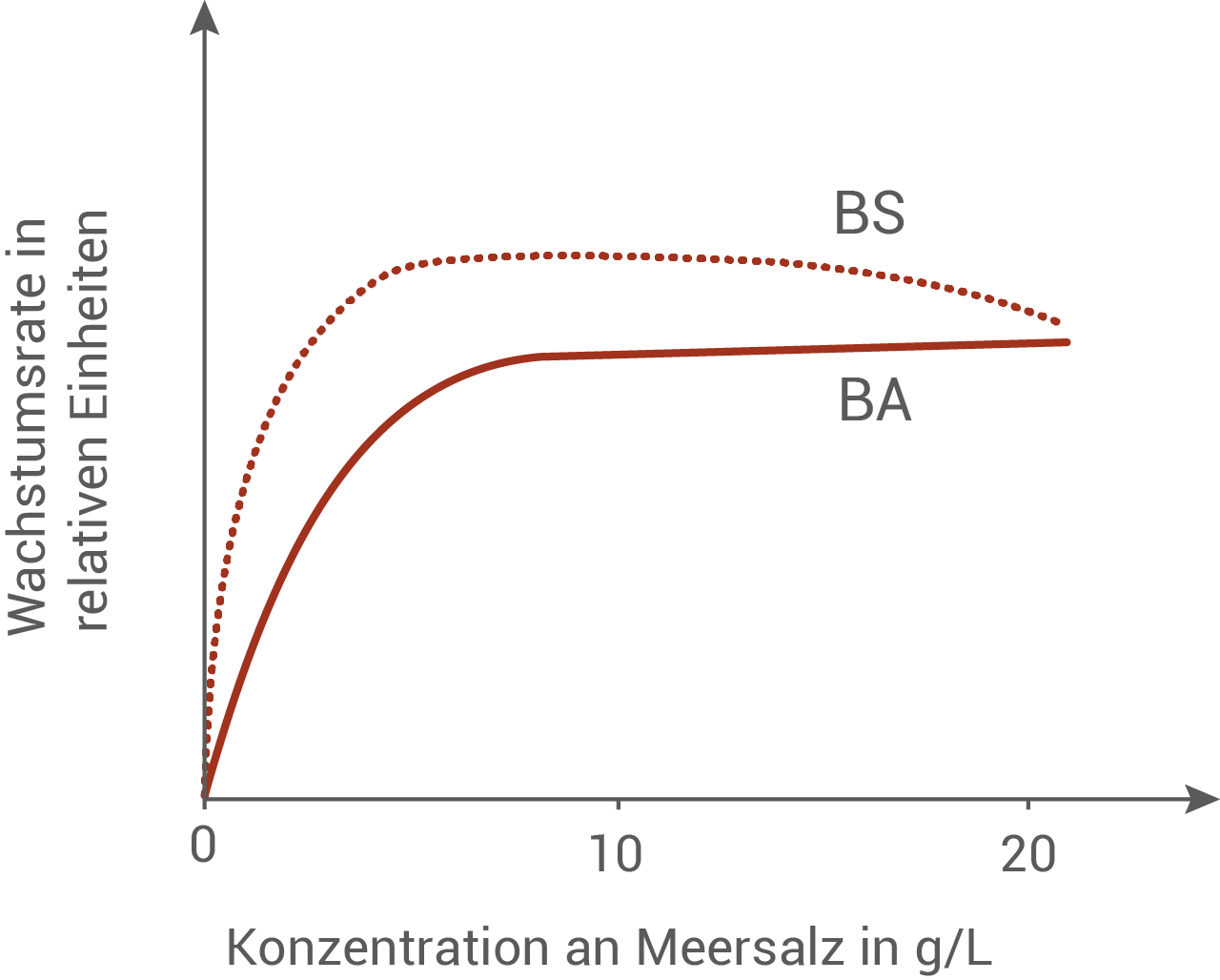
Beschreibe jeweils die Abhängigkeit der Wachstumsrate von der Meersalzkonzentration für beide Populationen von S. marinoi.
Erläutere das Zustandekommen der Unterschiede in der Salztoleranz der verschiedenen Populationen von S. marinoi mithilfe der erweiterten Evolutionstheorie.
Beschreibe ein molekularbiologisches Verfahren, mit dem die Verwandtschaftsverhältnisse der verschiedenen Populationen von S. marinoi ermittelt werden können.
In einem weiteren Versuch wurde das Fortpflanzungsverhalten von S. marinoi in Abhängigkeit von unterschiedlichen Außenfaktoren untersucht. Die Alge pflanzt sich ungeschlechtlich fort. Steigt jedoch der Salzgehalt im Wasser schnell stark an, pflanzt sie sich auch geschlechtlich fort.
Erkläre den Wechsel von ungeschlechtlicher Fortpflanzung zu einer geschlechtlichen Fortpflanzung unter diesen veränderten Umweltbedingungen aus ultimater Sicht.
Die im Plankton vorkommende einzellige Alge der Art Braarudosphaera bigelowii enthält ein besonderes Zellorganell, den Nitroplasten, durch den die Zelle in der Lage ist, Stickstoff zu fixieren. Ursprünglich war dieses Zellorganell ein prokaryotisches Cyanobakterium der Art Atelocyanobacterium thalassa.
Nenne zwei Gemeinsamkeiten und zwei Unterschiede zwischen Pro- und Eukaryoten.
In Abbildung 3 ist der Verlauf einer ungeschlechtlichen Vermehrung von B. bigelowii durch Zellteilung schematisch dargestellt.
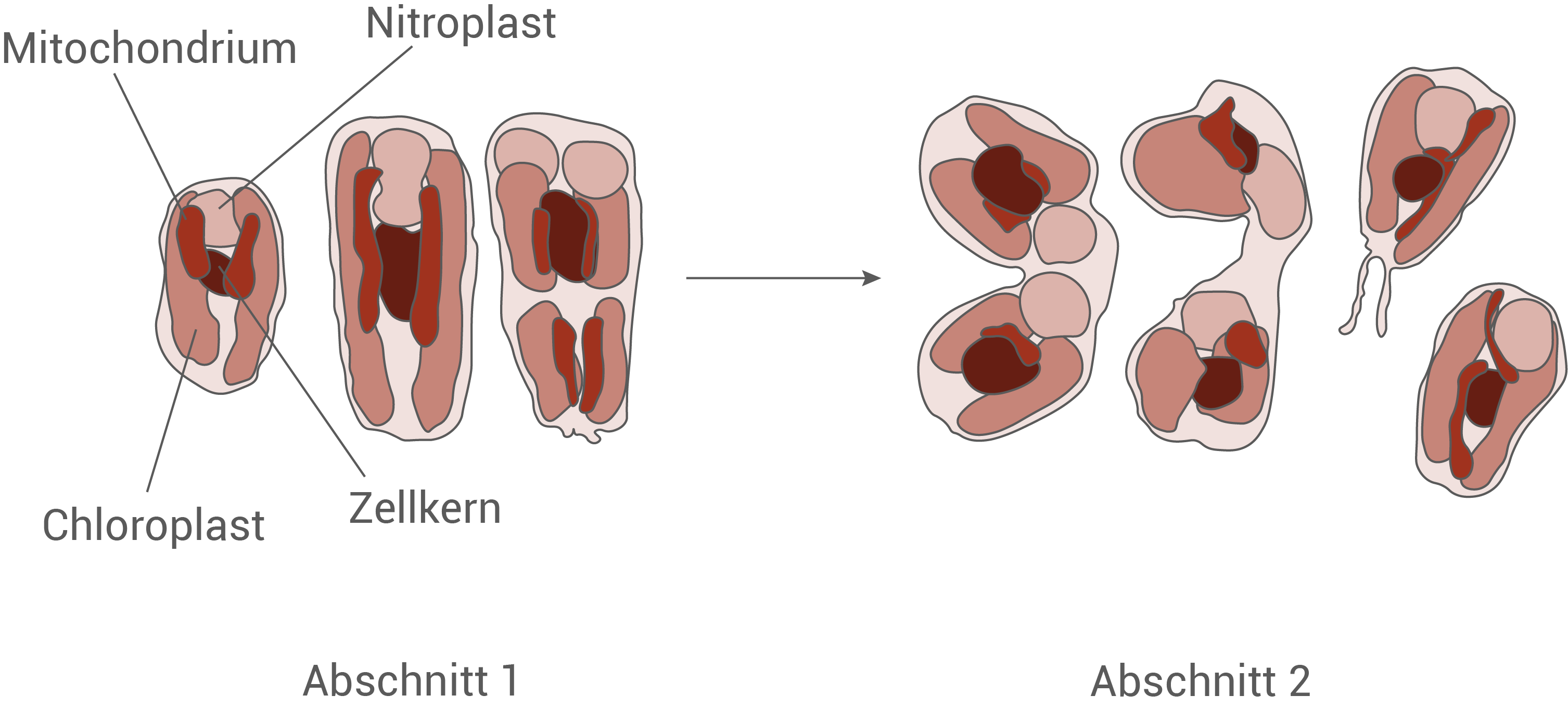
Beschreibe mit Hilfe der Abbildung 3 den Ablauf der Zellteilung der einzelligen Alge B. bigelowii und erläutere die Phasen der Mitose, die zwischen Abschnitt 1 und Abschnitt 2 ablaufen.
Bei bestimmten Dinoflagellaten, den Warnowiaceae, handelt es sich um eine Familie von einzelligen Eukaryoten des Planktons, die lichtempfindliche Strukturen aufweisen. Abbildung 4 zeigt einen Ausschnitt des Stammbaums dieser Familie. In Abbildung 5 sind verschiedene lichtempfindliche Strukturen dieser Dinoflagellaten dargestellt.
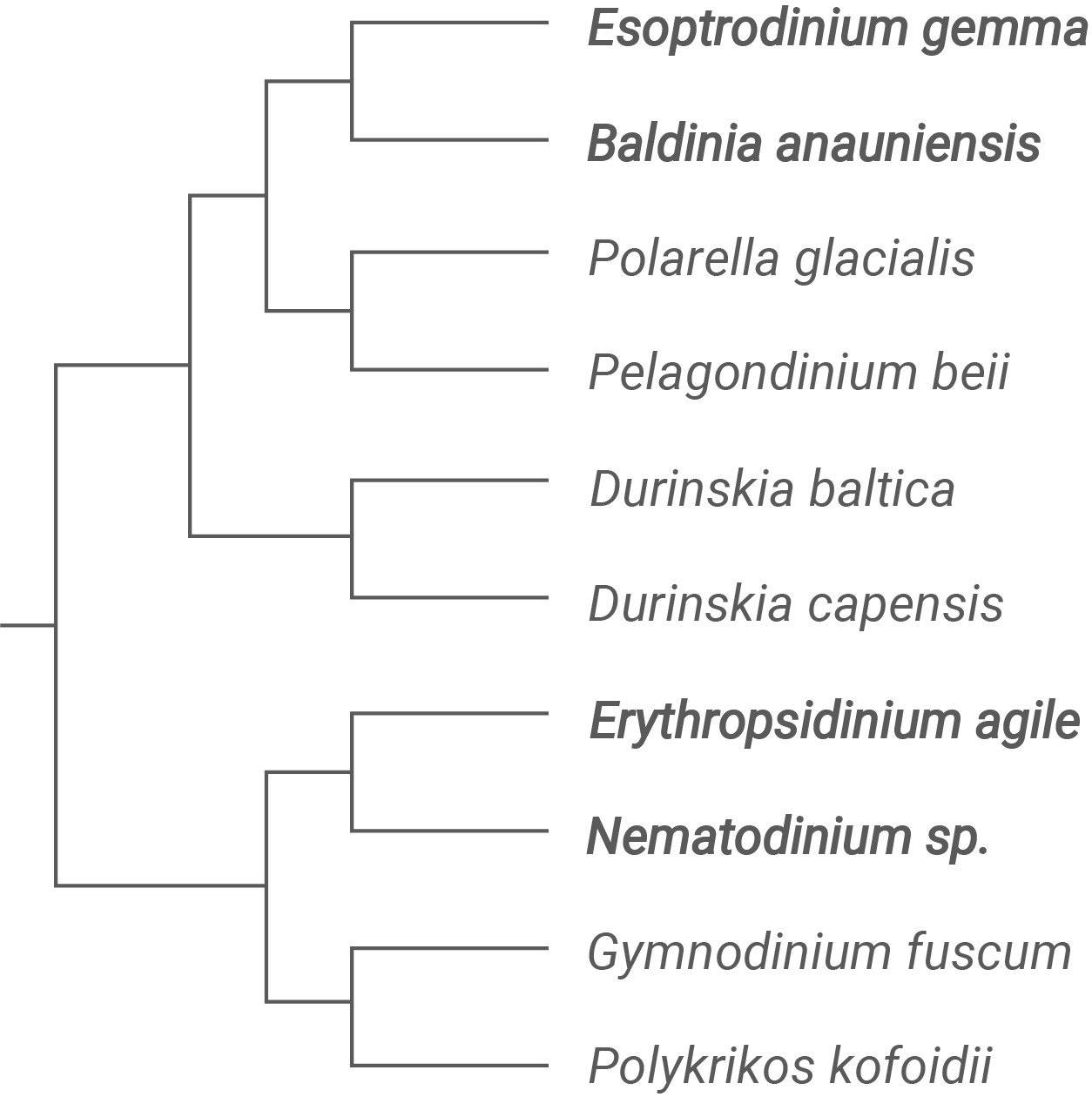

Diskutiere anhand von Abbildung 4 und 5, inwieweit die drei Homologiekriterien erfüllt sind, um eine Homologie der lichtempfindlichen Strukturen von B. anauniensis und Nematodinium sp. zu belegen.
Weiter lernen mit SchulLV-PLUS!
monatlich kündbarSchulLV-PLUS-Vorteile im ÜberblickDu hast bereits einen Account?Wachstumsrate für Populationen von S. marinoi je Meersalzkonzentration
Für die Population an der Stelle BS zeigt die Wachstumsrate bereits bei niedrigen Salzkonzentrationen vergleichsweise hohe Werte (starke Steigung). In diesem unteren Konzentrationsbereich bei einer Konzentration von etwa 5 g/L liegt ihr Optimum, also die maximale Wachstumsrate. Die Meersalzkonzentration an Stelle BS liegt, damit übereinstimmend, tatsächlich auch bei 5 g/L. Mit weiter zunehmender Meersalzkonzentration nimmt die Wachstumsrate jedoch kontinuierlich und immer stärker ab. Die Population der Kieselalge ist also perfekt an die Salzkonzentration ihres Standorts angepasst. Positive Abweichungen in der Konzentration verträgt sie tendenziell besser als negative, dennoch sinkt die Wachstumsrate auch bei hohen Salzkonzentrationen immer stärker.
Für die Population an der Stelle BA nimmt die Wachstumsrate mit steigender Meersalzkonzentration zunächst bis zu einer Konzentration von ungefähr 6 g/L auch deutlich zu. Der Wachstumsanstieg ist weniger stark und erreicht an keiner Stelle die gleiche Höhe wie die Population an der Stelle BS bei je gleicher Salzkonzentration. Im Gegensatz zur anderen Population sinkt die Wachstumsrate jedoch bei steigenden Konzentrationen nicht ab sondern steigt langsam, aber kontinuierlich an. Die Konzentration an Meersalz an der Stelle BA liegt bei 15 g/L. Die entsprechende Population zeigt bei dieser Konzentration eine relativ hohe Wachstumsrate, die aber weder am Maximum liegt noch bei höheren Konzentrationen abfällt, wie es bei der Population an der Stelle BS der Fall ist. Die Population bei BA zeigt also eine Anpassung an einen breiteren Toleranzbereich der Meersalzkonzentration (von etwa 6 g/L bis über 20 g/L) mit ähnlicher Wachstumsrate, aber dennoch der Tendenz zu einer höheren Wachstumsrate bei höheren Konzentrationen.
Erklärung der unterschiedlichen Salztoleranz
Die in Abbildung 2 erkennbaren Unterschiede in der Salztoleranz der beiden Populationen von Skeletonema marinoi lassen sich mit der erweiterten Evolutionstheorie als Ergebnis lokaler Anpassung an unterschiedliche Umweltbedingungen erklären. Die Population an der Stelle BA lebt langfristig in einem Lebensraum mit hohen Meersalzkonzentrationen, die Population bei BS aus der Ostsee dagegen in einem deutlich salzärmeren Milieu. Der Salzgehalt wirkt hier als abiotischer Selektionsfaktor, der die Überlebens- und Fortpflanzungschancen der Mikroalgen entscheidend beeinflusst, entsprechend deren Wachstumsraten.
In beiden Regionen treten in jeder Generation durch Mutation und Rekombination geringfügig unterschiedliche Genotypen auf, die sich in ihrer Salztoleranz unterscheiden. Individuen, deren genetische Ausstattung ein besonders gutes Wachstum bei den jeweils vorherrschenden Salzgehalten ermöglicht, besitzen eine höhere reproduktive Fitness: Sie teilen sich häufiger und tragen überproportional viele ihrer Allele in den Genpool der jeweiligen Population ein. Dies führt über viele Generationen hinweg zu einer gerichteten Selektion, bei der in der Population an BA solche Allele angereichert werden, die ein Optimum des Wachstums bei hohen Salzkonzentrationen bewirken, während in der Population an BS vor allem genetische Varianten überleben, die ein gutes Wachstum bei niedrigen Salzgehalten ermöglichen. So kommt es zu einer genetischen Differenzierung der Populationen entlang des Salzgradienten.
Hinzu kommt, dass der Genfluss zwischen beiden Regionen durch ozeanographische Bedingungen eingeschränkt ist, sodass sich die Allelfrequenzen der Populationen relativ unabhängig voneinander entwickeln können. Geringer Austausch begünstigt, dass sich lokale Anpassungen stabil ausprägen. Ergänzend berücksichtigt die erweiterte Evolutionstheorie neben genetischer Variation auch phänotypische Plastizität und epigenetische Regulation: Unterschiedliche Salzgehalte können über veränderte Genexpression kurzfristig Anpassungsreaktionen auslösen. Werden solche regulativen Fähigkeiten mit der Zeit genetisch abgesichert und selektiv begünstigt, verstärkt dies die Anpassung an die jeweilige ökologische Nische. Insgesamt entstehen so zwei Populationen derselben Art, die aufgrund unterschiedlicher Selektionsdrücke und begrenzten Genflusses unterschiedliche Wachstumsoptima bei verschiedenen Meersalzkonzentrationen ausgebildet haben.
Verwandtschaftsverhältnisse molekularbiologisch ermitteln
Hinweis: Zur Erreichung der vollen Punktzahl reicht es aus, ein einziges molekularbiologisches Verfahren zu beschreiben, was sich zur Prüfung der Verwandtschaftsverhältnisse eignet. Im Folgenden sind einige Beispiele solcher Verfahren erläutert.
DNA-Sequenzierung
Zur Bestimmung der Verwandtschaftsverhältnisse zwischen den beiden Populationen von Skeletonema marinoi kann nach der DNA-Isolation eine PCR durchgeführt werden, um bestimmte genetische Marker gezielt zu vervielfältigen. Dabei werden kurze einzelsträngige DNA-Stücke, sogenannte Primer, verwendet, die spezifisch an den Anfang und das Ende des zu untersuchenden DNA-Abschnitts binden. Die PCR läuft in wiederkehrenden Zyklen ab, bestehend aus Denaturierung, Annealing und Elongation. Während der Denaturierung trennt die hohe Temperatur die doppelsträngige DNA in zwei Einzelstränge. In der Annealing-Phase hybridisieren die Primer bei abgesenkter Temperatur an ihre komplementären Zielsequenzen. In der Elongationsphase synthetisiert die hitzestabile DNA-Polymerase, meist die Taq-Polymerase, neue DNA-Stränge, indem sie Nukleotide in 5'-3'-Richtung ergänzt. Durch die Wiederholung dieser drei Schritte entstehen schließlich Millionen Kopien des gleichen DNA-Abschnitts, die für eine präzise Sequenzbestimmung zur Verfügung stehen.
Anschließend kann die Basensequenz der vervielfältigten DNA mithilfe der Sanger-Sequenzierung ermittelt werden. Dabei werden zusätzlich zu normalen Nukleotiden im Puffer auch wenige modifizierte Nukleotide, sogenannte Didesoxynukleotide (ddNTPs), für den Ansatz eingesetzt. Diese ddNTPs besitzen keine 3'-OH-Gruppe und führen deshalb beim zufälligen Einbau während der Synthese zum Abbruch der DNA-Verlängerung. In der Sequenzierungsreaktion entstehen dadurch viele unterschiedlich lange DNA-Fragmente, die jeweils mit einem ddNTP enden. Die ddNTPs können fluoreszent markiert werden, wodurch nach der Trennung der Fragmente mittels Kapillarelektrophorese bestimmt werden kann, welches Nukleotid an welcher Position eingebaut wurde. Die geordnete Abfolge der Fluoreszenzsignale ermöglicht die exakte Rekonstruktion der Basensequenz des untersuchten DNA-Abschnitts. Auf diese Weise lassen sich die Sequenzen der beiden Populationen direkt vergleichen und daraus Rückschlüsse auf ihre genetische Verwandtschaft ziehen.
DNA-Hybridisierung
Bei der DNA-Hybridisierung wird ein Teil der doppelsträngigen DNA beider Populationen zunächst in einzelsträngige Form überführt. Einzelstränge der zu vergleichenden Populationen werden zusammengeführt und es wird geprüft, in welchem Ausmaß sie wieder miteinander hybridisieren, also komplementäre Basenpaare ausbilden. Je stärker die genetischen Sequenzen übereinstimmen, desto stabiler und weitreichender bilden sich Doppelstränge zwischen den Populationen aus. Die Stabilität der Hybridisierung lässt sich über die Schmelztemperatur bestimmen: Eine hohe Schmelztemperatur zeigt eine hohe Basenpaarungsrate und damit eine enge Verwandtschaft an. Eine geringe Hybridisierung oder niedrige Stabilität weist auf genetische Unterschiede hin. Die DNA-Hybridisierung erlaubt somit eine schnelle und vergleichsweise kostengünstige Abschätzung der genetischen Nähe zwischen den Populationen anhand der Messung der Schmelzpunktabsenkung zwischen reiner DNA und hybrid DNA.
Aminosequenzbestimmung von Proteinen
Verwandtschaftsbeziehungen können indirekt auch über den Vergleich der Aminosäuresequenzen homologer Proteine ermittelt werden. Zunächst wird ein repräsentatives Protein, etwa ein Enzym aus dem Fotosynthese- oder Stoffwechselapparat, aus beiden Algenpopulationen isoliert und gereinigt. Anschließend erfolgt eine Sequenzbestimmung der Aminosäurekette, entweder über Edman-Abbau oder moderne massenspektrometrische Verfahren. Da Proteine direkt von DNA-Sequenzen codiert werden, spiegeln Sequenzabweichungen im Protein Unterschiede in der zugrunde liegenden DNA wider. Je ähnlicher die Aminosäuresequenz zweier Populationen ist, desto enger sind sie miteinander verwandt.
Wechsel des Fortpflanzungsverhaltens
In der Verhaltens- und Evolutionsbiologie beschreibt die ultimative Sicht die evolutionsbiologischen Gründe und Fitnessvorteile eines Merkmals oder Verhaltens. Es geht also nicht um die unmittelbaren, physiologischen Mechanismen (das wären proximative Ursachen), sondern darum, warum ein Verhalten langfristig durch natürliche Selektion begünstigt wurde und den Fortpflanzungserfolg einer Art steigert.
Bei Skeletonema marinoi erfolgt unter stabilen Umweltbedingungen mit gleichbleibendem Salzgehalt die ungeschlechtliche Fortpflanzung, die eine schnelle Zellteilung und damit eine effiziente Vermehrung innerhalb kurzer Zeit ermöglicht. Die ungeschlechtliche Fortpflanzung maximiert somit die Populationsgröße, solange die Umweltbedingungen weitgehend konstant bleiben und keine tiefgreifenden Selektionsereignisse zu erwarten sind. In solchen Situationen ist genetische Variation weniger wichtig als schnelles Wachstum und Ressourcennutzung.
Steigt der Salzgehalt jedoch plötzlich und stark an, bedeutet dies einen massiven Umweltstress, der eine schnelle Anpassungsfähigkeit erfordert. Unter solchen Bedingungen entsteht durch ungeschlechtliche Vermehrung keine genetische Vielfalt, sodass alle Nachkommen identische genetische Eigenschaften besitzen und gemeinsam einem erhöhten Sterberisiko ausgesetzt sind. Die geschlechtliche Fortpflanzung erzeugt hingegen genetische Rekombination, wodurch neue Genotypkombinationen entstehen, die sich in ihrer Salztoleranz unterscheiden können. In einer variablen oder extremen Umwelt erhöht diese genetische Variation die Wahrscheinlichkeit, dass wenigstens ein Teil der Population überlebt. Die geschlechtliche Fortpflanzung wirkt also als evolutionäre Anpassungsstrategie, die langfristig den Fortpflanzungserfolg und damit die Fitness der Art sichert.
Pro- und Eukaryoten
Prokaryoten und Eukaryoten weisen zunächst einige grundlegende Gemeinsamkeiten auf. Beide besitzen eine Zellmembran, die den Zellinnenraum nach außen abgrenzt und den Stoffaustausch reguliert. Außerdem enthalten sowohl prokaryotische als auch eukaryotische Zellen Erbmaterial in Form von DNA, das die genetische Information für Aufbau und Funktion der Zelle trägt und bei der Zellteilung weitergegeben wird.
Daneben gibt es jedoch wichtige Unterschiede. Prokaryoten besitzen keinen von einer Membran umschlossenen Zellkern, ihre DNA liegt frei im Cytoplasma. Eukaryoten dagegen verfügen über einen Zellkern, in dem die DNA durch eine Kernhülle vom Cytoplasma getrennt ist. Ein weiterer grundlegender Unterschied betrifft die Zellorganellen. Eukaryotische Zellen besitzen verschiedene membranumhüllte Organellen wie Mitochondrien, Plastiden oder das endoplasmatische Retikulum, in denen bestimmte Stoffwechselprozesse räumlich getrennt ablaufen. Prokaryoten verfügen über solche membranbegrenzten Organellen nicht, ihre Stoffwechselprozesse finden überwiegend direkt im Cytoplasma oder an der Zellmembran statt.
Hinweis: Zur Erreichung der vollen Punktzahl können auch andere Gemeinsamkeiten bzw. Unterschiede herausgestellt werden.
Ungeschlechtliche Vermehrung durch Zellteilung
In Abbildung 3 ist die ungeschlechtliche Vermehrung von Braarudosphaera bigelowii als Zellteilung dargestellt. In Abschnitt 1 sieht man eine einzelne Mutterzelle mit einem Zellkern und den typischen Zellorganellen, darunter unter anderem Chloroplasten, Mitochondrien und den Nitroplasten. Im Verlauf der Zellteilung verdoppeln sich diese Zellorganellen in Abschnitt 1, indem sie sich jeweils durch eigene Teilung vermehren, sodass kurz vor der eigentlichen Teilung der Zelle jede Organelle mindestens in zweifacher Ausführung vorliegt. Auf diese Weise steht für jede der späteren Tochterzellen ein vollständiger Satz an Zellorganellen zur Verfügung. Zwischen Abschnitt 1 und Abschnitt 2 durchläuft der Zellkern eine Mitose. In Abschnitt 2 erkennt man schließlich zwei Tochterzellen, die jeweils einen Zellkern und einen eigenen Satz der zuvor genannten Organellen besitzen, wodurch nach der dargestellten Cytokinese zwei genetisch weitgehend identische Zellen entstehen.
Die Mitose des Zellkerns, die zwischen Abschnitt 1 und Abschnitt 2 abläuft, gliedert sich in die typischen Phasen Prophase, Metaphase, Anaphase und Telophase. In der Prophase kondensiert das chromosomale Erbmaterial, das zuvor als lockeres Chromatin im Zellkern vorlag, zu sichtbar unterscheidbaren Chromosomen, die bereits aus zwei Schwesterchromatiden bestehen. Die Kernhülle beginnt sich aufzulösen und es bildet sich der Spindelapparat aus Mikrotubuli, der von den Zentrosomen beziehungsweise Spindelpolen ausgeht. In der Metaphase ordnen sich die Chromosomen mit ihren Zentromeren in der Äquatorialebene der Zelle an, der sogenannten Metaphasenplatte. Die Spindelfasern sind nun mit den Zentromeren verbunden und stellen sicher, dass jede Chromatide über eine Spindelfaser mit einem der beiden Zellpole verknüpft ist. In der Anaphase trennen sich die Schwesterchromatiden am Zentromer und werden als nun eigenständige Tochterchromosomen aktiv entlang der Spindelfasern zu den entgegengesetzten Polen der Zelle gezogen. Dadurch erhält jeder Pol einen vollständigen Chromosomensatz. In der Telophase bilden sich um die an den Polen angekommenen Chromosomensätze neue Kernhüllen, die Chromosomen entspiralieren wieder zu lockerem Chromatin, und der Spindelapparat wird abgebaut. Darauf folgt abschließend die in Abschnitt 2 dargestellte Cytokinese.
Homologie zwischen zwei Dinoflagellaten
Zunächst ist zu klären, was mit den drei Homologiekriterien gemeint ist. Das Lagekriterium prüft, ob Strukturen im gleichen Bezug zum Gesamtbauplan des Lebewesens (bzw. dessen „Körper“) auftreten. Das Kriterium der spezifischen Qualität fragt nach besonderen Übereinstimmungen im Feinbau, die über bloße Funktionsähnlichkeit hinausgehen. Das Kontinuitätskriterium betrachtet, ob sich eine Struktur stammesgeschichtlich über Zwischenformen und den Stammbaum hinweg nachvollziehen lässt. Auf dieser Grundlage lässt sich nun diskutieren, inwieweit die lichtempfindlichen Strukturen von B. anauniensis und Nematodinium sp. als homolog gelten können. Grundlage sind der Stammbaum der Warnowiaceae in Abbildung 4 und die schematischen Zeichnungen der lichtempfindlichen Strukturen in Abbildung 5.
Beim Lagekriterium zeigen die Abbildungen zunächst deutliche Übereinstimmungen. Sowohl bei B. anauniensis als auch bei Nematodinium sp. liegt die lichtempfindliche Struktur in einem ähnlichen Bereich der Zelle, nämlich peripher in der Zellrinde und in räumlicher Nähe zum Flagellenapparat beziehungsweise in Bewegungsrichtung der Zelle. In beiden Zeichnungen ist sie klar als eigener Komplex erkennbar und nicht irgendwo diffus im Cytoplasma verteilt. Die relative Position im Gesamtbauplan des Dinoflagellaten ist also vergleichbar, was das Lagekriterium weitgehend erfüllt.
Beim Kriterium der spezifischen Qualität fallen ebenfalls deutliche Parallelen auf, aber auch Unterschiede. In Abbildung 5 zeigt Nematodinium sp. ein deutlich komplexes ocelloidartiges Organell mit klar abgrenzbarer linsenartiger Struktur, einem retinaähnlichen lichtempfindlichen Bereich und einer deutlichen membranösen Umschließung. Diese Merkmale wirken stark differenziert und erinnern in ihrer funktionellen Anordnung an ein augenähnliches System, das gerichtete Lichtwahrnehmung erlaubt. Auch bei B. anauniensis ist eine lichtempfindliche Struktur erkennbar, die dieselben Grundfunktionen besitzt, jedoch in einem wesentlich einfacheren und weniger differenzierten Aufbau. Zwar lassen sich auch hier mindestens eine lichtbündelnde Region und ein lichtempfindlicher Bereich erkennen, doch wirken Größe, Ausprägung und Abgrenzung weniger deutlich differenziert als beim komplexen Ocelloid von Nematodinium sp.. Gerade diese qualitative Abstufung legt zunächst nahe, dass beide Organellen auf ein gemeinsames Grundmuster zurückgehen und sich im Verlauf der Evolution unterschiedlich stark spezialisiert haben. Das Organell von B. anauniensis ließe sich damit als vereinfachte Variante derselben lichtempfindlichen Struktur interpretieren, während Nematodinium sp. eine stark weiterentwickelte Form besitzt. Aus rein morphologischer Sicht spricht diese abgestufte Komplexität dafür, dass beide Strukturen nicht unabhängig entstanden sind, sondern Varianten einer gemeinsamen Ausgangsform darstellen könnten.
Beim Kontinuitätskriterium kann man zuerst Abbildung 5 hinzuziehen: Die in dargestellten, unterschiedlich komplexe, lichtempfindliche Strukturen verschiedener Arten der Familie erweckt den Eindruck, dass es sich um eine Progressionsreihe handeln könnte. Dies würde unterstreichen, dass es sich durch den Merkmalszusammenhang über diese unterschiedlich komplexen Zwischenstufen um ein homologes Merkmal handelt. Schwieriger wird die Bewertung bei Einbezug des Stammbaums in Abbildung 4. Die Abbildung zeigt B. anauniensis und Nematodinium sp. innerhalb der Familie Warnowiaceae, sie sitzen aber nicht direkt nebeneinander, sondern sind durch mehrere andere Gattungen getrennt. Lichtempfindliche Strukturen treten nicht bei allen dargestellten Taxa gleichermaßen auf, manche besitzen komplexe „Augenorganelle“, bei anderen sind diese Strukturen deutlich einfacher. Zusätzlich lässt sich aufgrund der frühen Aufspaltung der Arten Esoptrodinium gemma und Baldinia anauniensis von den Arten Erythropsidinium agile und Nematodinium sp. nicht eindeutig feststellen, dass die lichtempfindlichen Strukturen über die Bildung neuer Arten tatsächlich im Laufe einer Progressionsreihe zunehmend komplexer wurden.. Daraus ergeben sich zwei konkurrierende Lesarten. Entweder man nimmt an, dass ein ocelloidartiges Organ einmal im gemeinsamen Vorfahren der Warnowiaceae entstanden ist, im Stammbaum dann aber mehrfach reduziert oder entwickelter wurde. In diesem Fall wäre die Kontinuität über den Stammbaum hinweg prinzipiell gegeben, die großen Abstände müssten jedoch über zahlreiche Modifikationen und Verluste erklärt werden. Alternativ kann man annehmen, dass sich in verschiedenen Linien der Warnowiaceae, also auch unabhängig in der Linie von B. anauniensis und von Nematodinium sp., ähnliche lichtempfindliche Strukturen konvergent entwickelt haben, weil die selektiven Anforderungen an gerichtete Lichtwahrnehmung vergleichbar waren. In diesem Fall wäre das Kontinuitätskriterium nur teilweise erfüllt, da es keine klare, lückenlose Verteilung einer gemeinsamen Vorläuferstruktur im Stammbaum gibt. Die relativ große phylogenetische Distanz der beiden Arten spricht eher dafür, hier vorsichtig zu argumentieren und eine hundertprozentig belastbare Kontinuitätslinie zu bezweifeln. In der Gesamtschau ergibt sich daher ein differenziertes Bild. Das Lagekriterium und die spezifische Qualität der lichtempfindlichen Strukturen sprechen durchaus dafür, dass B. anauniensis und Nematodinium sp. auf ein gemeinsames, augenähnliches Grundorganell in der Familie der Warnowiaceae zurückgehen könnten. Das Kontinuitätskriterium ist dagegen durch die große Distanz im Stammbaum und die uneinheitliche Verteilung ähnlicher Strukturen in der Gruppe nur eingeschränkt erfüllt. Damit ist eine Homologie der lichtempfindlichen Strukturen beider Arten zwar gut begründbar, aber nicht zweifelsfrei belegt. Genauso vertretbar ist die Interpretation, dass die ocelloidartigen Organelle innerhalb der Warnowiaceae aus ähnlichen Bausteinen konvergent entstanden sind. Für eine abiturechte Diskussion bedeutet das: Zwei Homologiekriterien stützen eher die Annahme einer Homologie, das dritte liefert durch die phylogenetische Situation gewichtige Einwände, sodass man im Fazit von einer plausiblen, aber nicht eindeutig gesicherten Homologie sprechen sollte.